- 昆虫の変態を制御する遺伝子を操作して、カメムシが成虫の姿形への変化を早めたり失わせたりすることに成功
- カメムシの変態に伴って、腸内共生細菌を保持する器官が食物消化の機能も持つようになる原因、また、腸内共生細菌が卵殻の構成成分を生産してカメムシの旺盛な繁殖力を支えることを解明
- 変態や共生の機構を標的とした害虫制御法の開発に貢献が期待される

チャバネアオカメムシの変態を制御する遺伝子を操作して、成虫になるタイミングを失った個体(左から2番目)と 通常の脱皮回数より早く成虫になった個体(左から3番目)を作り出した
国立研究開発法人 産業技術総合研究所(以下「産総研」という)生物プロセス研究部門 生物共生進化機構研究グループ森山 実 主任研究員、二橋 亮 上級主任研究員、深津 武馬 首席研究員(東京大学大学院理学系研究科 教授(兼任))は、東京大学大学院理学系研究科生物科学専攻博士後期課程(当時)の大石 紗友美 元産総研技術研修員、水谷 雅希 日本学術振興会特別研究員と、腸内共生細菌が生存に必要不可欠なチャバネアオカメムシにおいて、(i) 変態前の幼虫と変態後の成虫で、消化管の後端部に発達する共生器官の形態と機能が異なること、(ii) 共生器官の幼虫型から成虫型への切り替えが変態制御遺伝子でコントロールされていること、(iii) 幼虫型の共生器官は腸内共生細菌の保持に特化しているのに対して、成虫型の共生器官はそれに加えて食物の消化吸収を行うようになること、(iv) 成虫の腸内共生細菌は卵殻形成に必要なアミノ酸を多量に合成するようになること、(v) これら変態に伴うカメムシ自身と腸内共生細菌それぞれの変化が、多量の食物を摂取して数日ごとに卵塊を産むカメムシ成虫の旺盛な繁殖力を支えていることを解明しました。
本研究により、昆虫類の多様性と繁栄を支える要因の一つである変態が、昆虫自身のみならず、腸内共生細菌の機能も制御していることが判明しました。共生関係にある異種生物間の高度な機能的統合を明らかにした重要な成果であり、変態や共生の機構を標的とした害虫制御法の開発に寄与する可能性もあります。
なお、この研究成果の詳細は、2023年9月25日の週(米国東部時間時間)に米国の学術誌「Proceedings of the National Academy of Sciences of the United States of America」(米国科学アカデミー紀要)にオンライン掲載されます。
地球上に満ちあふれる生物は、生態系の中で互いに影響を及ぼし合いながら、それぞれの生息環境に適応し、進化してきました。そのようにして生じた高度な生物機能の理解は、基礎生物学的に興味深いのみならず、さまざまな形で応用利用されてきました。
カメムシ類は世界に40,000種以上、日本には1,500種以上が分布し、多くの農業害虫を含みます。産総研生物プロセス研究部門では、昆虫類における高度な生物機能の解明に取り組んでおり、特に微生物との共生関係に着目してさまざまな研究を展開しています。カメムシ類の生存には特定の腸内共生細菌が必要で、これまでに「大腸菌を昆虫共生細菌に進化させることに成功」(2022年8月4日 産総研プレス発表1)などの研究成果を発表しました。一方で昆虫類の多様性や環境適応に深く関わる「変態」のメカニズムについても研究し、これまでに「トンボの幼虫から成虫への変態に必須な遺伝子群の同定に成功」(2022年2月22日 産総研プレス発表2)などを発表しました。しかし、昆虫の変態がどのように微生物との共生関係に関わるのかはよくわかっていませんでした。今回、チャバネアオカメムシの腸内共生系をモデルとして共生と変態の関係の解明に取り組み、新たな知見を得ることができました。
なお、本研究開発は、国立研究開発法人 科学技術振興機構「ERATO深津共生進化機構プロジェクト」および科研費(新学術領域研究等)による支援を受けて実施しました。
チャバネアオカメムシの消化管の後端は共生器官と呼ばれ、盲のうという袋状の構造が管に沿って4列に並び、その内部に生存に必要不可欠な細菌(腸内細菌科Pantoea 属の1種)を保持します(図1A)。先行研究において、幼虫と成虫で共生器官の構造が顕著に異なることが報告されています。幼虫では、共生器官の前端が完全に閉じていて、共生器官に食物が流入することはなく、食物は消化管の前半部で全て吸収されると考えられます(図1B-D)。一方、成虫では共生器官の前端は開口しており、食物が消化管から共生器官に流入しますが、盲のうの基部は閉鎖しています(図1E-G)。これらの違いはどのようなしくみで生じるのでしょうか。
昆虫類では、幼若ホルモンと脱皮ホルモンの作用によって、変態制御遺伝子の発現が変化して、幼虫から成虫への変態が起こります。具体的には、幼若ホルモンの存在下で幼虫化遺伝子(Kr-h1)が発現し、その状態で脱皮ホルモンが働くと、幼虫のまま変態しない脱皮が起こります。一方、幼若ホルモンがない状態では成虫化遺伝子(E93)が発現するようになり、その状態で脱皮ホルモンが働くと、チョウやハエやカブトムシなどではさなぎへの脱皮が、カメムシやトンボやバッタなどでは成虫への脱皮が起こります。
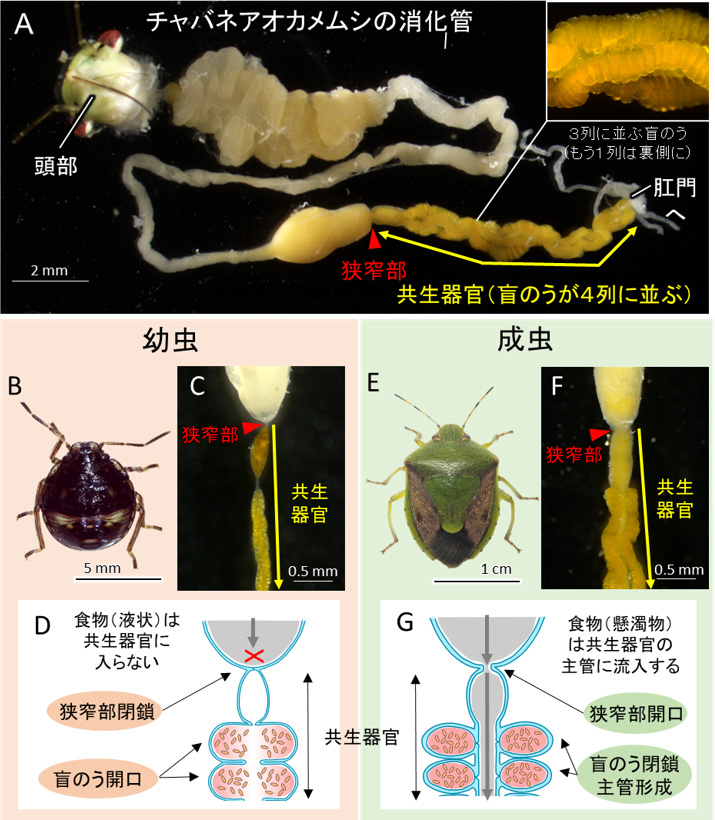
図1 チャバネアオカメムシの共生器官
(A)チャバネアオカメムシの摘出消化管。消化管後端の共生器官を黄矢印、共生器官の前端に位置する狭窄(きょうさく)部を赤矢頭で示す。枠内は共生器官の拡大像で盲のうの配列を示す。(B-D)幼虫およびその共生器官の写真および模式図。(E-G)成虫およびその共生器官の写真および模式図。
チャバネアオカメムシでは通常、5回目の脱皮を控えた幼虫(5齢幼虫)が脱皮して成虫になります(図2A、B)。ところがRNA干渉法という技術を用いて、5齢幼虫で成虫化遺伝子の発現を抑制したところ、脱皮した個体は幼虫の姿をした6齢の過齢幼虫になりました(図2C)。一方、4齢幼虫で幼虫化遺伝子の発現を抑制したところ、脱皮した個体は成虫の姿をした早熟成虫になりました(図2D)。これらの個体の共生器官を調べたところ、過齢幼虫では幼虫型の共生器官を、早熟成虫では成虫型の共生器官を持つことがわかりました(図2C、D)。6齢に成長して体が大きくなっても、成虫化遺伝子の発現が抑えられれば姿形も体内の共生器官も幼虫型になる一方で、5齢で若く体が小さくても、幼虫化遺伝子の発現が抑えられれば姿形のみならず体内の共生器官も成虫型になることが明らかになりました。加えて、共生器官における遺伝子発現パターンを解析した結果、食物が流入するようになった成虫型の共生器官は、実際に消化機能を担うことが示されました。すなわち、共生器官が幼虫型になるか成虫型になるかを決めているのは、発生段階(5齢か6齢か)でも成長の程度(体が大きいか小さいか)でもなく、変態制御遺伝子であることが示されました。
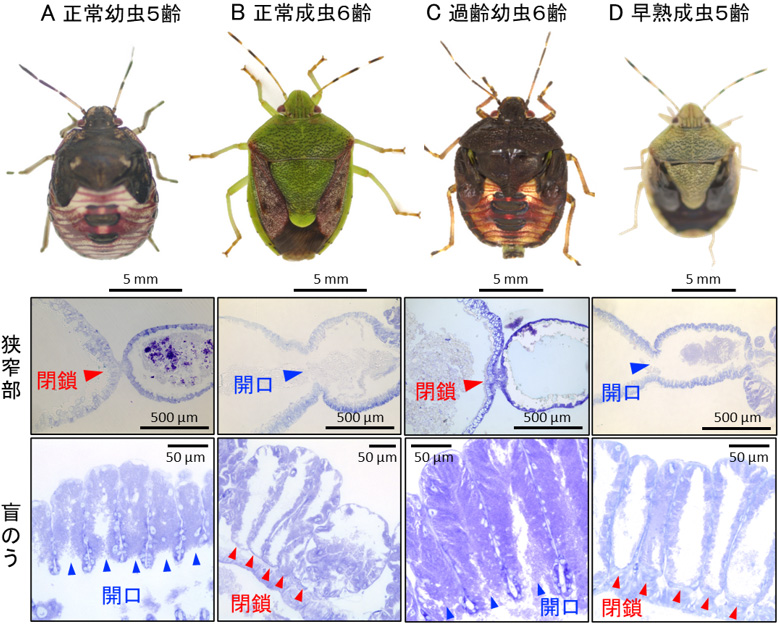
図2 チャバネアオカメムシ過齢幼虫および早熟成虫の共生器官の構造
(A)正常幼虫(5齢)。(B)正常成虫(6齢)。(C)過齢幼虫(6齢)。(D)早熟成虫(5齢)。上段は虫の外観、中段は狭窄部の組織像、下段は盲のうの組織像を示す。過齢幼虫は幼虫型の共生器官を、早熟成虫は成虫型の共生器官を持つことがわかる。
さらに、腸内共生細菌の遺伝子発現パターンを調べたところ、成虫の共生器官ではシステインなどのアミノ酸合成に関わる遺伝子の発現が軒並み顕著に増加していました。また、成虫体液中のシステイン含量は幼虫よりも有意に高くなっていました。カメムシにおけるシステインの含有量を調べたところ、餌のピーナツや成虫の体全体よりも卵に多く含まれ(図3B)、そのほとんどが卵殻に含まれていることがわかりました(図3C)。システインはジスルフィド結合を形成することでタンパク質の分子間を架橋し、構造的に強固にする働きがあり、硬くて丈夫な卵殻を形成するのに重要と思われます。チャバネアオカメムシのメス成虫は、十分に食物を与えると約14卵からなる卵塊を2〜3日に1個、1カ月以上にわたり産み続けるという旺盛な繁殖力を示しますが(図3A)、成虫カメムシの腸内共生細菌は、その繁殖力を支えるためにシステインを高生産するよう制御されているものと考えられます。
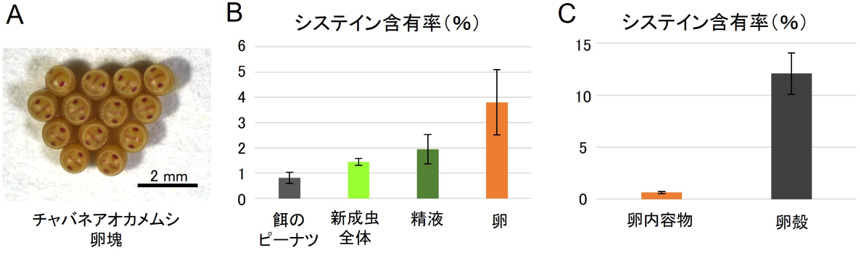
図3 チャバネアオカメムシ卵殻における高いシステイン含有率
(A)チャバネアオカメムシの卵塊。(B)餌のピーナツ、新成虫全体、オスの精液、メスの卵のシステイン含有率。(C)卵の内容物および卵殻のシステイン含有率。
これらの研究から、チャバネアオカメムシが幼虫から成虫になる際に、体内の共生器官も変態制御遺伝子によって幼虫型から成虫型へ形態と機能が変化すること、さらに腸内共生細菌も幼虫型から成虫型へ機能が変化することが明らかになりました。これらの変化は、共生器官および腸内共生細菌の機能が、幼虫ではもっぱら自身の生育を支える共生関係の維持であるのに対して、成虫では旺盛な摂食のための食物消化および産卵のための卵殻成分の合成に切り替わるという、「幼虫期における成長」と「成虫期における繁殖」という生育段階に伴う機能的分業が基盤になっており、生物学的に意義深いことです。本研究により、昆虫と微生物という共生関係にある生物の間で、「変態」という劇的な発生過程をめぐり、高度な機能的統合が行われている実態が示されました。
本研究で、変態制御遺伝子として知られる幼虫化遺伝子(Kr-h1)および成虫化遺伝子(E93)により、共生器官および腸内共生細菌が幼虫型になるか成虫型になるか決まることを明らかにしましたが、具体的な実行部隊の遺伝子はKr-h1やE93に制御される下流遺伝子と考えられるため、それらの解明に引き続き取り組んでいきます。
また、変態をつかさどる幼若ホルモンや脱皮ホルモンが、宿主昆虫の共生器官の形態や機能を制御する具体的なしくみも今後の研究課題です。特に、腸内共生細菌がこれらのホルモンに直接的に制御されているとは考えにくく、共生器官における何らかの変化から間接的な制御を受けているものと思われますが、その実体の解明にも取り組んでいきます。
カメムシ類を含む農業害虫種の変態を撹乱する薬剤は、すでに農薬として利用されています。昆虫の生存に必須な腸内共生細菌1や、昆虫の殺虫剤抵抗性に関わる腸内共生細菌3も、害虫制御の新規標的として期待されています。昆虫における変態と共生の関係の理解は、これらの害虫制御技術の開発にも貢献すると考えられます。
掲載誌:Proceedings of the National Academy of Sciences of the United States of America
論文タイトル:Regulation and remodeling of microbial symbiosis in insect metamorphosis
著者:Sayumi Oishi, Minoru Moriyama, Masaki Mizutani, Ryo Futahashi, Takema Fukatsu