国立研究開発法人産業技術総合研究所(以下「産総研」という)健康医工学研究部門 三村 真大 技術研修生(研究当時)、石原 紗綾夏 テクニカルスタッフ、栗田 僚二 研究部門付、冨田 峻介 研究グループ長、人工知能研究センター 亀田 倫史 上級主任研究員、細胞分子工学研究部門 新海 陽一 研究グループ長らは、国立大学法人東京科学大学 総合研究院 自律システム材料学研究センター 菅井 祥加 特任助教、立命館大学 薬学部 北原 亮 教授、北沢 創一郎 助教(研究当時)と共同で、液-液相分離が遺伝子の働きを制御する仕組みに着目し、ヒストンタンパク質とDNAからなる液滴の生じやすさが、ヒストンの化学修飾を受ける部位によって変化することを明らかにしました。
私たちの生命機能は、どの遺伝子がいつ・どこで働くかによって制御されています。この制御には、転写因子やエピジェネティクスなど、多様な仕組みが関わっており、特にDNAが巻き付くヒストンに施される化学修飾は、代表的な調節機構として広く研究されてきました。近年では、細胞内でタンパク質やDNAが自発的に集まり、周囲とは区別された“液滴”状の構造をつくる「液-液相分離(liquid-liquid phase separation: LLPS)」という現象が知られるようになり、こうした液滴が、特定の分子を集めたり隔離したりすることで、遺伝子の働きを調節する場として機能する可能性があると考えられています。しかし、ヒストンの化学修飾と液-液相分離がどう関連するのかは十分に明らかではありませんでした。また、こうした制御機構の異常は、がんや神経変性疾患などとの関連も指摘されています。
研究グループはこれまで、DNAとタンパク質による液-液相分離の分子機構とその機能の解明に取り組んできました。今回、ヒストンの化学修飾の一つであるアセチル化に着目し、ヒストンのどの部位がアセチル化されるかによって、ヒストンとDNAからなる液滴の生じやすさが異なることを明らかにしました。これは、ヒストンの化学修飾の「有無」だけでなく、修飾される「場所」が液滴形成を制御することを示す成果であり、液滴形成を介した遺伝子制御の理解に新たな視点を与えるものです。今後、遺伝子の発現異常を標的とした創薬や診断技術の開発に向けた新たな手掛かりとなることが期待されます。
なお、この技術の詳細は、2026年5月7日に「Journal of the American Chemical Society」に掲載されました。
がんや神経変性疾患(アルツハイマー病など)の多くは、細胞内での遺伝子発現制御の異常が関与すると考えられています。そのため、遺伝子の働きがどのように制御されているかを分子レベルで理解することは、新たな治療法や創薬標的の発見につながると期待され、世界中で研究が進められています。
遺伝子発現の制御には、DNAの配列を変えずにその働きを調節するエピジェネティクスが重要な役割を果たしており、その代表的な仕組みの一つが、DNAを巻き付けるヒストンに生じる化学修飾(翻訳後修飾)です。中でも、ヒストンのアセチル化は、がんや神経変性疾患と深く関わっていることが知られています。
一方、近年では、細胞内のタンパク質やDNAが自発的に集まり、水と油が自然と分かれるように“液滴”状の構造形成が誘導される液-液相分離という現象が、遺伝子制御の仕組みの一つとして注目されています。液滴は、遺伝子の働きに関わる分子を集めたり隔離したりすることで、発現の制御に関与すると考えられていますが、その制御の変化がさまざまな疾患と関連する可能性も指摘されています。
産総研では、生体分子がどのように相互作用して液滴を形成し、その結果としてどのような生命機能が生じるのかを、分子レベルで理解することを目指し、研究を進めてきました。これまでに、DNAが形成する特殊な立体構造であるグアニン四重鎖がヒストンとの相互作用を通じて液滴形成を促進することを明らかにし、DNAの構造が液-液相分離の制御に関わることを示してきました[1-3]。
今回の研究では、この流れをさらに発展させ、DNA側の構造に加えて、ヒストン側の化学修飾に着目しました。特にヒストンの代表的な修飾であるアセチル化について、修飾部位の違いが液滴形成に与える影響を明らかにしました。これにより、産総研がこれまで進めてきた「DNA/ヒストン相互作用による相分離制御」の研究を、エピジェネティクスの観点へと拡張することができました。
なお、本研究開発は、独立行政法人日本学術振興会 科学研究費助成事業 基盤研究(B)(2023~2025年度)および特別研究員奨励費(2020~2021年度)、国立研究開発法人日本医療研究開発機構の委託事業 脳神経科学統合プログラム(精神・神経疾患メカニズム解明プロジェクト)「相分離破綻に起因する神経変性疾患に関する研究開発」(2021~2024年度)による支援を受けています。
細胞核の中では、DNAは8つのヒストン(H2A, H2B, H3, H4がそれぞれ2分子)に巻き付いてヌクレオソームという基本構造を作っています(図1A)。ヒストンにはDNAの外側へ伸びた「テール」と呼ばれる部分があり、このテールの化学修飾が遺伝子の制御に関わることが知られています。こうした修飾は、同じ種類であっても、どの部位に入るかによって異なる生物学的役割を担うことが知られています。本研究では、この4種類のヒストンのうちH3のテール部分と、ヌクレオソーム間をつなぐリンカーDNAを模した二重鎖DNAからなるモデル実験系を構築し(図1B)、ヒストンのアセチル化の「有無」だけでなく、どの「場所」がアセチル化されるかによって液滴形成のしやすさがどう変化するかを、実験とシミュレーションを組み合わせて解析しました。
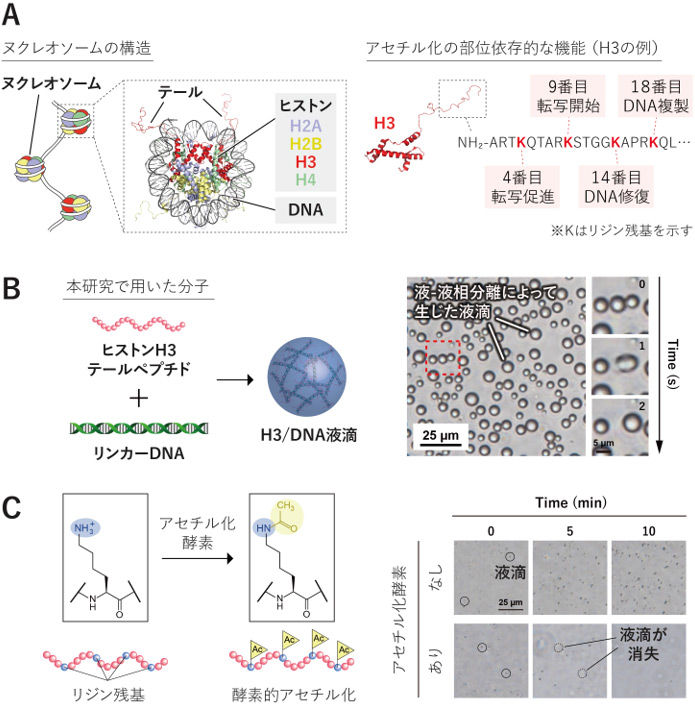
図1 本研究で着目したヒストンH3テールのアセチル化部位と、H3/DNA液滴形成の概要
A:DNAはヒストンに巻き付いてヌクレオソームという構造を形成します。ヒストンにはDNAの外側に伸びたテール(ヒストンテール)と呼ばれる部位があり、ここには複数のアセチル化部位が存在します。H3テールのアセチル化部位は、それぞれ異なる生物学的機能と関わることが知られています。ヌクレオソームの立体構造には、ヒストンテールを含む代表的な既知構造として、アフリカツメガエル由来の構造(PDB: 1KX5)を示しました。B:本研究では、ヒストンH3のテールの一部を化学合成したペプチド(ヒストンH3テールペプチド)とリンカーDNAを模した二重鎖DNAを用いたモデル系を構築しました。両者を混合すると液-液相分離により液滴が形成され、液滴どうしが融合する様子が観察されました。C:H3テール中のリジン残基はアセチル化酵素によりアセチル化されます。酵素を加えると、形成されていた液滴は時間とともに消失し、H3のアセチル化が液滴形成を抑制することが示されました。
※原論文の図を引用・改変したものを使用しています。
まず、ヒストンH3のテールの一部を化学合成したペプチド(以下、H3ペプチド)とリンカーDNAを模した35塩基対の二重鎖DNAを混合すると、球状の液滴が形成され、液滴どうしが融合する様子が観察されました(図1B)。ここに、ヒストンをアセチル化する酵素を加えると、形成されていた液滴が時間とともに消失しました(図1C)。つまり、H3ペプチドとDNAの組み合わせは液-液相分離によって液滴を形成し、その形成はアセチル化によって抑制されることがわかりました。
次に、アセチル化される部位の違いが液滴形成に与える影響を調べるため、特定のリジン残基だけを選択的にアセチル化したH3ペプチドを化学合成しました(図2)。その結果、アセチル化は全体として液滴形成を抑制する一方で、その抑制の強さは修飾部位によって異なることがわかりました。具体的には、H3ペプチドの中央付近がアセチル化された場合には液滴形成が比較的保たれたのに対し、末端付近がアセチル化された場合には液滴形成が強く抑制されました(図2)。これにより、液滴形成のしやすさは、アセチル化の有無だけでなく、その部位によっても大きく左右されることが明らかになりました。このような液滴形成の違いが、遺伝子発現に関わる分子の集まり方や働き方に影響を与える可能性があると考えられます。
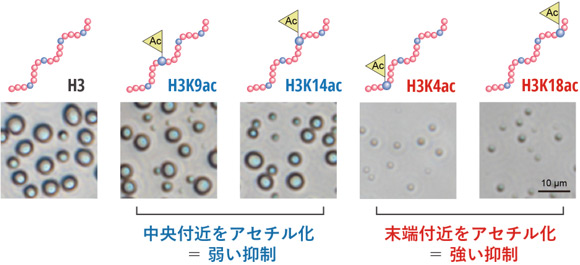
図2 ヒストンH3のアセチル化部位によって液滴形成の抑制の強さが異なる
未修飾のH3ペプチド、および特定のリジン残基のみをアセチル化したH3ペプチド(H3K9ac、H3K14ac、H3K4ac、H3K18ac)について、DNAと混合した際の液滴形成を比較しました。中央付近(K9、K14)がアセチル化された場合には液滴形成が比較的保たれたのに対し、末端付近(K4、K18)がアセチル化された場合には液滴形成が強く抑制されました。これにより、液滴形成のしやすさは、アセチル化の有無だけでなく、その部位によっても大きく左右されることが示されました。
※原論文の図を引用・改変したものを使用しています。
では、なぜヒストンテールのアセチル化部位によって液滴形成のしやすさが変わるのでしょうか。これを理解するため、全原子分子動力学シミュレーションを用いて、H3ペプチドがDNAのどこに結合しやすいかを調べました(図3)。その結果、未修飾H3ペプチドやテールの中央付近がアセチル化されたH3ペプチド(H3K9ac)では、ペプチドがDNAの両端付近に集まりやすいのに対し、テールの末端付近がアセチル化されたH3ペプチド(H3K4ac)では、ペプチドがDNA全体により均一に分布する傾向が観察されました。H3ペプチドは全体としてプラスの電荷を、DNAはマイナスの電荷を帯びているため、ペプチドの結合位置が偏るか均一かによって、H3ペプチド/DNA複合体の表面に生じる電荷分布も変化します。例えば、テールの末端付近のアセチル化では複合体表面の電荷がより均一になると考えられます。
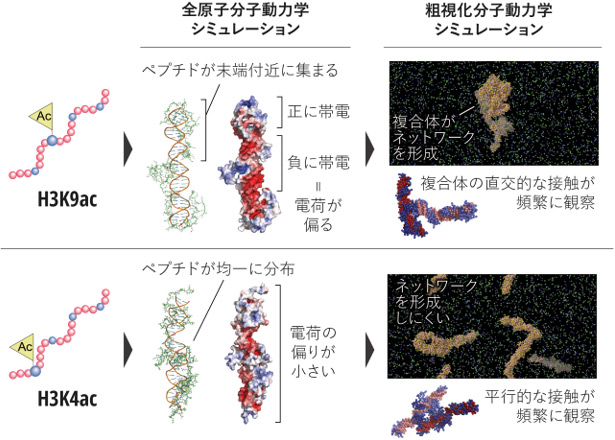
図3 分子動力学シミュレーションから示された、アセチル化部位による相互作用の違い
上段はテールの中央付近がアセチル化されたH3ペプチド(H3K9ac)、下段は末端付近がアセチル化されたH3K4acの結果を示しています。全原子分子動力学シミュレーションの結果、H3K9acではペプチドがDNAの末端付近に集まりやすく、複合体内で電荷の偏りが生じる一方、H3K4acではペプチドがDNA全体により均一に分布し、電荷の偏りが小さいことが示されました。さらに粗視化分子動力学シミュレーションでは、H3K9ac由来の複合体は直交的な接触をとりやすく、複合体間ネットワークを形成しやすいのに対し、H3K4ac由来の複合体は平行的な接触が多く、ネットワークを形成しにくいことが示されました。これらの結果は、アセチル化部位の違いが、複合体内の電荷分布と複合体間相互作用を変化させ、これが液滴形成のしやすさと関係する可能性を示唆しています。
※原論文の図を引用・改変したものを使用しています。
さらに、この複合体を用いて粗視化分子動力学シミュレーションを行ったところ、未修飾H3ペプチドやテールの中央付近がアセチル化されたH3ペプチドからなる複合体では、複合体どうしが直交的に接触しやすく、その結果、複数方向に連結したネットワーク構造が形成されやすいことが示唆されました。一方、テールの末端付近がアセチル化されたH3ペプチドからなる複合体では、平行に並ぶ接触が多く、連結の方向が限定されるため、ネットワークが形成されにくいことが示されました(図3)。前者のような接触が生じやすい理由としては、複合体の両端が比較的プラスに、中心部分が比較的マイナスに帯電しているため、異なる複合体どうしの間での静電的な引き合いによる橋渡しが起こりやすいことが考えられます。こうした複合体間ネットワークの形成しやすさの違いが、液滴形成の生じやすさに違いをもたらしているとみられます。
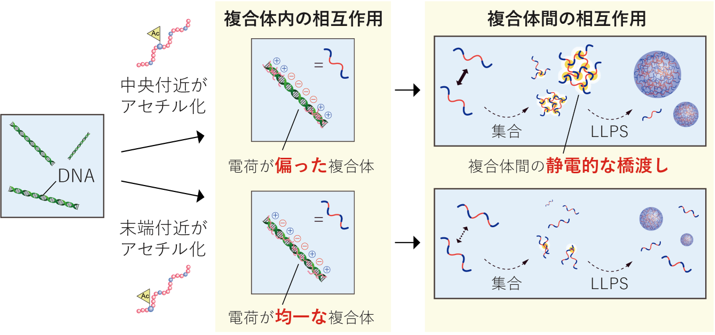
図4 ヒストンH3のアセチル化部位によって液滴形成のしやすさが変化する分子モデル
中央付近がアセチル化された場合、H3ペプチド/DNA複合体内では電荷の偏りが比較的保たれた構造が形成されます。その結果、複合体どうしの間で静電的な橋渡しが起こりやすく、複合体間ネットワークが形成されやすいため、液-液相分離が比較的維持されると考えられます。一方、末端付近がアセチル化された場合には、H3/DNA複合体内の電荷分布がより均一になり、複合体間ネットワークが形成されにくくなるため、液-液相分離(LLPS)が強く抑制されると考えられます。
※原論文の図を引用・改変したものを使用しています。
以上の知見を基に、本研究では、ヒストンのアセチル化部位が液滴形成を制御する分子モデルを提案しました(図4)。テールの中央付近がアセチル化されると、複合体内の電荷分布の偏りが比較的保たれ、静電的相互作用で複合体間ネットワークが形成されやすいため、液滴形成が比較的保たれます。一方、テールの末端付近がアセチル化されると、複合体がより均一な電荷分布をとり、複合体間ネットワークが形成されにくくなるため、液滴形成が強く抑制されると考えられます。本研究は、ヒストン修飾の部位情報が液滴形成を制御するという新たな分子基盤を提示するものです。
今後は、他のヒストンテールやさまざまな翻訳後修飾にも対象を広げ、液-液相分離を介した遺伝子制御の普遍的な仕組みの解明を目指します。併せて、異なる配列や長さのDNA、ヒストン結合タンパク質などを取り入れた、より生体に近い実験系へ展開することで、エピジェネティクスと液-液相分離の関係をより深く理解するための基盤技術の確立を目指します。さらに、今回得られた「修飾部位によって液滴形成が制御される」という知見をもとに、相分離異常やヒストン修飾異常を標的とした創薬研究や、クロマチン状態を制御する機能性分子の設計につながる基盤へと発展させていきます。
掲載誌:Journal of the American Chemical Society
論文タイトル:Proximity of the histone-acetylation site to the termini shapes phase behavior with DNA
著者:Masahiro Mimura, Hiroka Sugai, Tomoshi Kameda, Ryo Kitahara, Soichiro Kitazawa, Yoichi Shinkai, Sayaka Ishihara, Ryoji Kurita, and Shunsuke Tomita
DOI:https://doi.org/10.1021/jacs.6c02267
[1] Mimura, M. et al. Quadruplex Folding Promotes the Condensation of Linker Histones and DNAs via Liquid–Liquid Phase Separation. Journal of the American Chemical Society. 2021, vol. 143, no. 26, p. 9849-9857. DOI: 10.1021/jacs.1c03447.
[2] Sugai, H. et al. Emergence of Anisotropic Subcompartments via Coassembly of Hierarchically Ordered G-Quadruplexes and Fluid Polylysine in Droplet-Based Compartments. ACS Nano.2026, vol. 20, no. 13, p. 10520-10531. DOI: 10.1021/acsnano.5c21437.
[3] 産総研マガジン. DNAの立体構造と“液-液相分離”の関係を初めて証明―神経変性疾患の予防・治療につなげる(https://www.aist.go.jp/aist_j/magazine/20250829.html)