国立研究開発法人 産業技術総合研究所【理事長 中鉢 良治】(以下「産総研」という)生物プロセス研究部門【研究部門長 田村 具博】 植物機能制御研究グループ 坂本 真吾 産総研特別研究員、光田 展隆 主任研究員らは、国立研究開発法人 森林総合研究所【理事長 沢田 治雄】(以下「森林総研」という)森林バイオ研究センター【センター長 吉田 和正】森林バイオ研究室 高田 直樹 主任研究員、谷口 亨 室長と共同で、イネの遺伝子を使ってポプラの木質を大幅に強化する技術を開発した。
この技術はイネの木質生産を制御しているOsSWN1転写因子を、遺伝子組換えによりポプラに導入して、ポプラの成長には悪影響を及ぼさずに木質生産性を約4割高め、木材の強度も約6割向上させることができる。将来的には、木質由来のバイオエタノールやバイオプラスチックの高効率生産、高い強度を持った木材の開発、さらに木質由来の次世代燃料・材料の高効率生産がもたらすCO2の排出削減への貢献が期待される。
なお、本技術開発は、国立研究開発法人 科学技術振興機構(JST)戦略的創造研究推進事業(先端的低炭素化技術開発:ALCA)の研究開発課題の一環として行われ、2016年1月27日19時(日本時間)にScientific Reportsにオンライン掲載される。
 |
|
イネの木質生産を制御する転写因子遺伝子をポプラに導入して木質を増強 |
地球温暖化を抑制するため、また、将来枯渇する化石燃料を代替するため、植物由来の燃料や材料の開発が進められている。ところが現在の植物由来バイオエタノールやバイオマテリアルは、食糧生産との競合が問題となっており、食糧ではない木質を原料とした第二世代のバイオエタノールやバイオマテリアルの開発と普及が期待されている。このため、植物の木質生産性の向上は重要な課題の一つであるが、これまで木質生産を増強しようとすると植物の成長に悪影響が生じるといった問題があり、植物の成長を阻害せずに木質生産性を向上させる技術が求められていた。
産総研では、植物の転写因子に関する研究開発を進めており、これまでも独自の転写因子改変技術であるCRES-T法の開発や、それを利用した「バラ咲きシクラメン」の開発(2010年3月16日 産総研プレス発表)、クチクラ形成を制御する転写因子の発見(2013年5月24日 産総研プレス発表)、シロイヌナズナの木質生産を制御する転写因子NST1、NST3の発見などの世界をリードする成果を上げてきた。また、森林総研では樹木の育種や遺伝子組換え技術の開発で卓越した成果を挙げてきた。今回、両者は転写因子を利用して植物の木質の生産性や加工性を向上させる研究開発に取組み、本研究成果をあげることに成功した。
なお、この研究は、JSTの戦略的創造研究推進事業(先端的低炭素化技術開発:ALCA)の技術領域「バイオテクノロジー」【運営総括:近藤 昭彦】 研究開発課題「ゼロから創製する新しい木質の開発(平成23年度~)」【研究開発代表者:光田 展隆】の一環として行われた。
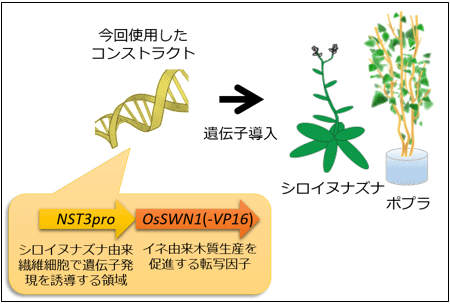
今回の技術は、シロイヌナズナの木質生産を制御するNST1転写因子と NST3転写因子の相同遺伝子であるイネの木質生産を制御するOsSWN1転写因子が、非常に強く木質生産を活性化できることを利用し、その遺伝子をポプラの繊維細胞で主に発現させるものである。このイネのOsSWN1遺伝子を繊維細胞で発現させるために、シロイヌナズナのNST3遺伝子の植物体内での発現部位を決定している領域(プロモーター)を使用した。NST3転写因子遺伝子は繊維細胞で主に発現しており、そのプロモーターは繊維細胞での遺伝子発現を誘導できる。これらを組み合わせた遺伝子コンストラクト(図1)を、シロイヌナズナに導入(遺伝子組換え)したところ、通常では木質生産が起きない部位でも木質生産が見られ、木質が過剰に蓄積することがわかった。一方、比較対照としてOsSWN1転写因子のかわりにシロイヌナズナが本来持つNST3転写因子を用いた場合は、そのような現象はほとんど見られなかった。
次にこの遺伝子コンストラクト中のイネOsSWN1遺伝子に、転写因子の活性を強化する領域(VP16)を付加して、ポプラに導入(遺伝子組換え)したところ、約15 cmの幼植物ではシロイヌナズナと同様に、通常では木質生産が起きない部位でも木質生産が見られた(図2B)。また、本来木質生産が起きる繊維細胞では、組換えポプラでは木質がより厚く蓄積していた(図2D)。
 |
|
図1 今回使用した遺伝子コンストラクト |
|
シロイヌナズナ由来繊維細胞で遺伝子発現を誘導する領域(NST3pro)とイネ由来木質生産を促進する転写因子遺伝子(OsSWN1(-VP16))とを連結した。 |
 |
|
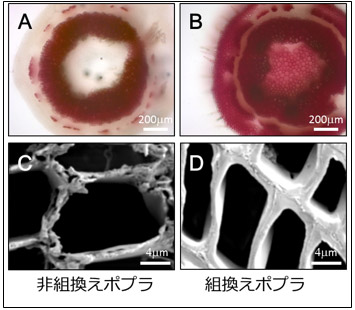
図2 非組換えポプラ(左側)と組換えポプラ(右側)の断面図と繊維細胞 |
|
AとBの赤い染色は木質中のリグニンの存在を示したもの。組換えポプラ(B)では通常木化しない部位(周縁部および中央部)まで木化していることがわかる。また、繊維細胞では、非組換えポプラ(C)より組換えポプラ(D)のほうが、より多く木質が蓄積していることが確認できる。 |
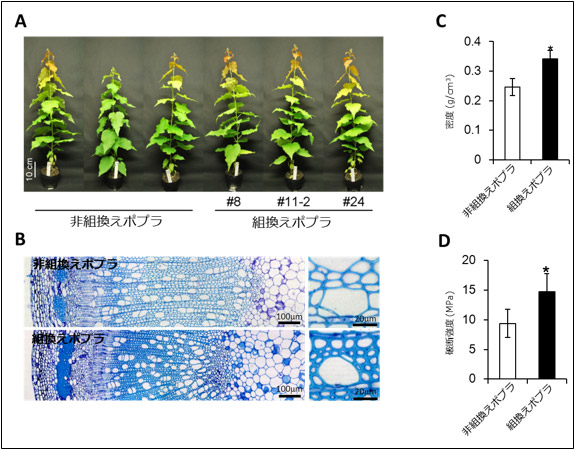
そこで、これらのポプラを約60 cmまで成長させて詳しく調べたところ、成長には悪影響を及ぼさず木質が過剰に蓄積し、組換えポプラの茎の平均密度は非組換えポプラよりも約4割向上し、破断強度も約6割上昇していた(図3)。
 |
|
図3 非組換えポプラと組換えポプラの生育状況(A)、木質(B)、密度(C)、破断強度(D) |
|
約60 cmまで生育させても成長に悪影響はなく(A)、木質の増強も維持されていることがわかる(B)。また、作成した組換えポプラの5系統の平均で約4割密度が向上し(C)、破断強度は約6割向上していた(D)。(C)(D)内の「*」は非組換えポプラと組換えポプラとが統計的に有意に異なった値であったことを示している。 |
今後は、光合成能力の強化など他のバイオマス生産向上技術と組み合わせて、さらなる生産量の増加を目指すほか、増強された木質中のリグニンを改変するなどして加工性や糖の抽出量の向上を目指す。また、ポプラだけでなく、ユーカリやアカシアなどの樹木への今回の技術の適用を検討する。そして、2030年頃には木質由来バイオエタノールの生産効率を50 %向上させ、全世界で栽培する木質生産用植物の20 %にこの技術を適用して、年間約4千万トンのCO2排出削減効果を得ることを目指す。