さまざまな生物の全ゲノム解読は、全遺伝子の解明を通じて広く生命科学に寄与するとともに、生物進化の研究に多くの知見をもたらしてきました。多くの動物は父方と母方からの同一のゲノムをもつ「二倍体」ですが、アフリカツメガエルは、異種交配と全ゲノム重複により一つの生物の中に異なる2種類のゲノムをもった「異質四倍体」とされていました。そのため、非常に有用なモデル生物であるにもかかわらず、全ゲノム解読が非常に困難と諦められ、主要モデル生物の中で唯一行われていませんでした。しかし日本とアメリカを中心とする国際コンソーシアムは、アフリカツメガエルの全ゲノム解読に挑み、見事その全貌を明らかにしました。得られた情報は今後生物学から医学に至るさまざまな研究分野に大きく貢献すると期待できます。加えて、アフリカツメガエルのゲノムの中にある2種類のゲノム(サブゲノム)が別々の染色体のセットに分かれて存在するという重要な発見をしました。それにより、このカエルは約1800万年前に、2つの種が異種交配と全ゲノム重複を起こして誕生した異質四倍体であること、その後2つのサブゲノムが一つの生物の中で異なる進化を辿ったことが明確に示されました。今日の地球上には実に多様な種類の脊椎動物が生息し繁栄していますが、その最大の要因と考えられるのが約5億年前の古生代カンブリア紀に起きたとされる「2回の全ゲノム重複」です。その謎を解くための重要な鍵、いわゆるロゼッタストーンとしてアフリカツメガエルのサブゲノムの進化の仕組みが役立つことになります。これは生命科学における画期的な成果です。
<背景と課題>
一つの生物がもつ全遺伝情報をゲノムと言い、その本体はDNAです。今日、種々の生物のゲノムDNAが解読されており、そこで得られたゲノム情報は生命科学の発展に大きく寄与しています。それと共にゲノム情報を生物間で比較することは、生物進化の研究に多くの知見をもたらしてくれます。それは、数十億年の生物の歴史のなかで途切れることなく子孫へと受け継がれてきたゲノムを調べれば、その中に痕跡として残されている進化過程を探し出すことができると考えられるからです。これまで脊椎動物のゲノム解読は、まずヒトで行われ、その後はマウスやゼブラフィッシュ、メダカなど世界的に多くの研究者に用いられている実験モデル生物を中心に行われてきました(図1)。アフリカツメガエル(図2)は、1950年代から現在に至るまで、動物の発生の仕組みや細胞の性質を調べる上で非常に有用な実験モデル動物として使われてきました。2012年に山中伸弥博士と共にノーベル生理学・医学賞を受賞したジョン・ガードン博士はこのカエルを用いて、「細胞の初期化」を初めて実験的に示したことで有名です。しかしながら、研究の歴史が古くこれまで多くの重要な発見をもたらしてきた主要モデル生物の中で、唯一ゲノム解読されていなかったのが、複雑なゲノムのため解読が困難とされていたアフリカツメガエルでした。
 |
|
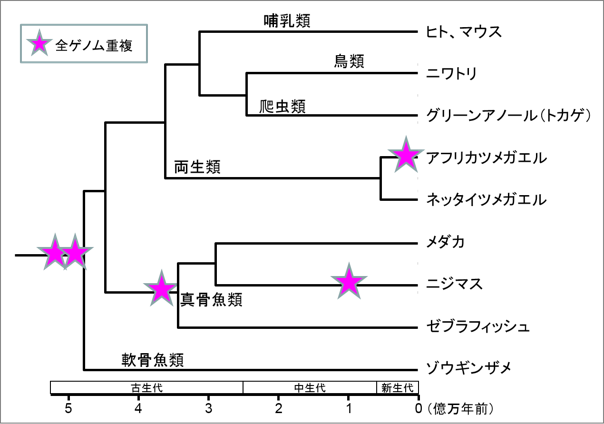
図1. 脊椎動物の系統樹と全ゲノム重複。系統樹は分類群の分岐年代に従って表し、右端にゲノム解読された動物名を示す。全ゲノム重複(星印)は、脊椎動物の共通祖先種で約5億年前に2回起きたとされている。さらに真骨魚類の共通祖先種では約3.2億年前に3回目の全ゲノム重複が起き、ニジマスの系統では1億年前にさらに4回目の全ゲノム重複が起きた。両生類ではアフリカツメガエルの系統で1800万年前に3回目の全ゲノム重複が起きた。 |
 |
|
図2. アフリカツメガエルとネッタイツメガエル。(a)成体メスの比較。外見は良く似ているが、アフリカツメガエルの方がネッタイツメガエルより大きい。(b)頭部の拡大図。アフリカツメガエル(上)とネッタイツメガエル(下)では顔つきが異なる。このアフリカツメガエルは近交系の J 系統である。(c)胚の比較。アフリカツメガエル(上)とネッタイツメガエル(下)の胚。アフリカツメガエルとネッタイツメガエルの卵の直径はそれぞれ1.2 mmと0.7 mmであり、アフリカツメガエルの方が大きく、この時期の胚も大きい。 |
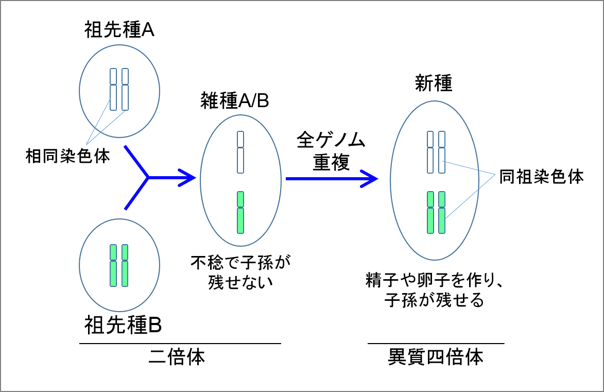
多くの生物は、父方と母方から受け継いだ同一種類のゲノムを2つもつ「二倍体」ですが、一つの生物の中に2種類のゲノムを2つずつもつものがあり、これを「異質四倍体」と言います(図3)。図3に示すように異質四倍体となるきっかけは近縁な2つの種の異種交配であり、そのあと染色体数の倍加、すなわち全ゲノム重複が起こり異質四倍体となります。アフリカツメガエルは、新生代の頃に2つの種の異種交配で生じた異質四倍体の種であると考えられていました。しかしこれら2つの祖先種は既に絶滅し、現存していません。このようにアフリカツメガエルのゲノムはいわば、1種のカエルの中に2種の絶滅した祖先種ガエルのゲノム(これをサブゲノムと言います;図4)が共存した状態といえます。したがって全ゲノム解読はこれらの2種類の互いに良く似たサブゲノムを区別して解読する必要があるため、非常にチャレンジングでありました。しかし、主要モデル生物として生命科学の発展に不可欠であること、また、脊椎動物の初期の進化の過程において起きたとされる2回の全ゲノム重複に重要な示唆を与えることから、2009年に日本と米国で期を同じくして独立にプロジェクトチームが立ち上がり、全ゲノム解読が始まりました。
 |
|
図3. 異質四倍体は雑種の全ゲノム重複によってつくられる。ここでは簡単にするため祖先種の染色体は1対のみを描いてある。実際の染色体数は、祖先種は9対もち、アフリカツメガエルは18対をもつ。 |
 |
|
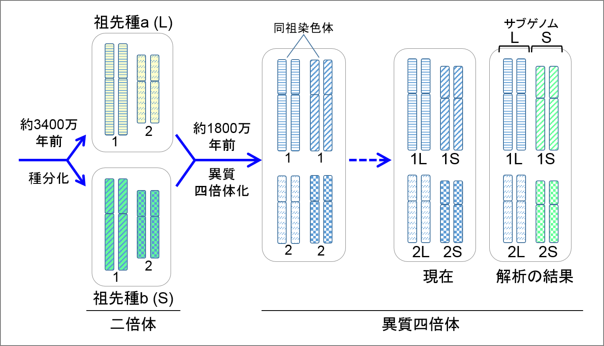
図4. 異質四倍体は祖先種に由来する2つのサブゲノムをもつ。ここでは簡単にするため祖先種 a と b の染色体は1番と2番の2対のみを描いた(実際は祖先種は9対でアフリカツメガエルは18対である)。異質四倍体化の直後は、同祖染色体間に区別がないが現在までに一方が短くなったと考えられる。そこで長い方を L(long)、短い方を S(short)と命名した。今回、詳細なゲノム解析を行った結果、染色体 L のセットと S のセットが、祖先種由来のゲノム(これをサブゲノムという)にそれぞれ対応することが示された。そこで、二つのサブゲノムを L と S と命名し、さらに祖先種も L と S と命名した。この発見により、倍数化後のサブゲノムの変化を解析することが可能となった。 |
<研究内容>
日本チーム(代表:東京大学・平良眞規)と米国チーム(代表:カリフォルニア大学・ダニエル・ロクサーとリチャード・ハーランド)は、2012年に国際コンソーシアムとして共同でゲノム解読を行うことで合意しました。それを可能にしたのが、日本が独自に作出した近交系動物(J 系統、図2)を、両チームが用いたことです。J 系統はゲノムのDNA塩基配列に個体差がないため、2種の祖先種由来のサブゲノムの塩基配列の違いを浮かび上がらせることができました。それにより米国チームは、短く断片化したDNAの塩基配列を明らかにし、それらをパズルのピースのようにつなげていくことが可能となりました。しかしそれだけではよく似た2つのサブゲノム由来の塩基配列を区別して解読するには不十分です。日本チームの国立遺伝学研究所の藤山秋佐夫・豊田敦グループは、非常に長いDNA断片の塩基配列を明らかにし、名古屋大学の松田洋一・宇野好宣グループがそれらのDNA断片がアフリカツメガエルの染色体18対のうちどの染色体に対応するかを何百も調べました。さらに日本ツメガエル研究会(XCIJ)を母体とする研究グループ(広島大学・鈴木厚、北海道大学・福井彰雅、長浜バイオ大学・荻野肇、東京大学・近藤真理子ら16名)が、根気の要る緻密な確認作業を丹念に行いました。これらの共同作業により、ようやくゲノムの全体を、しかも非常に正確に、染色体ごとに解読することに成功しました。
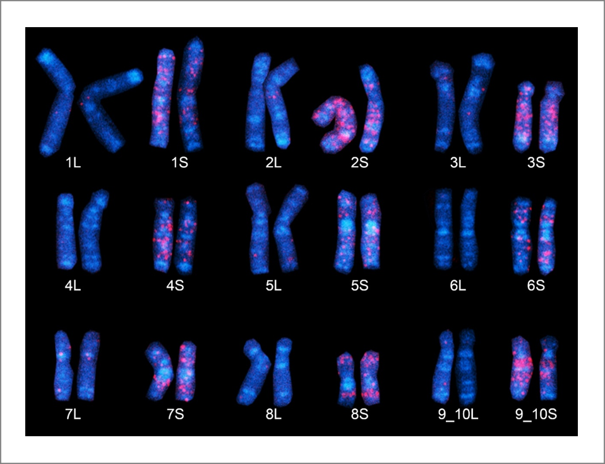
次に、解読された全ゲノムDNA塩基配列を用いた解析を行いました。広島大学の彦坂暁グループは、「化石化した」トランスポゾンのDNA塩基配列に注目することで、2つの祖先種から受け継いだサブゲノムをみごとに区別しました(図4と図5)。驚いたことに、2つのサブゲノムは一つの細胞の中でそれぞれが9本の染色体のセットとして維持されていました。つまり絶滅した祖先種それぞれがもっていた9本の染色体のセットがほぼそのままアフリカツメガエルの中に残っていたことになります。しかも注意深く比較すると、一方の染色体セットの染色体の長さが他方に比べて少しずつ短いことが分かりました(図4と図5)。そこで長い染色体のセットを L(long)、短い方を S(short)と名付け、それらの起源となる絶滅した祖先種も L と S、さらにそれに対応するサブゲノムも L と S と名付けました。
 |
|
図5. アフリカツメガエルのサブゲノムの同定。サブゲノム S に特異的な"化石化"DNA配列を用いて、染色体(青)をFISH法で赤く染色したもの。染色体1番~9番(9_10番)の S の染色体により多くの赤い染色が見られる。これらは祖先種 S に由来したものと考えられる。9番目の染色体は、ネッタイツメガエルの9番と10番染色体が融合した染色体に相当するため染色体9_10番と呼ぶ。 |
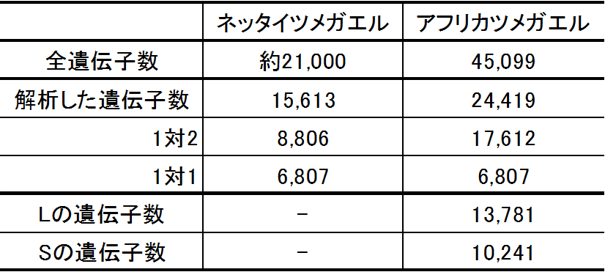
サブゲノム L と S が区別できたことで、2つの祖先種が誕生したのが約3400万年前であること、それらが異種交配して異質四倍体になったのが、新生代の中新世に入った約1800万年前であることが分かりました(図4)。アフリカツメガエルの遺伝子はゲノム中に全部で45,099個見つかりました。この数は二倍体の近縁の種のネッタイツメガエルの約2倍でした(表1)。染色体に存在する遺伝子を対応させると、ネッタイツメガエルの1本に対してアフリカツメガエルの2本の染色体 L と S が丁度対応しました。そこでさらに詳しく比較をすると、染色体セット L の方がネッタイツメガエルの染色体に良く似ており、染色体セット S の方がより多くの遺伝子が無くなっていることが分かりました。さらに使われ方にも大きな差があり、染色体セット L に存在する遺伝子の方がより多く使われていました。これらの結果から、異質四倍体になる時の全ゲノム重複の後、どのようにサブゲノムが進化するかが初めて明らかになりました。
 |
|
表1 遺伝子数の比較と同祖遺伝子の保持について。 |
<結論と今後の展望>
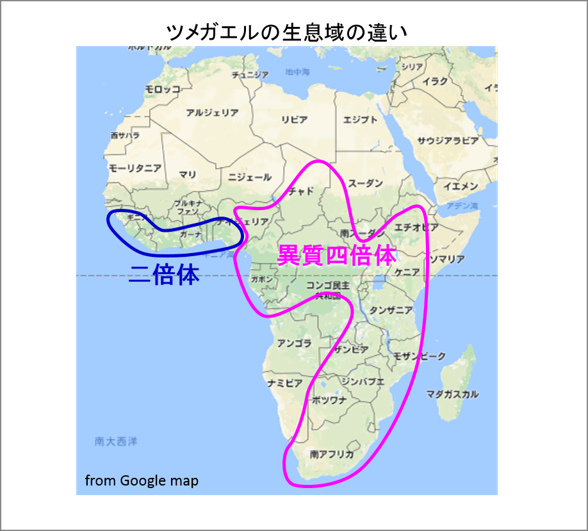
では異質四倍体になることの利点は何だったのでしょうか。ツメガエル属のカエルの生息域を見てみますと、二倍体の種は赤道付近に限られていますが、異質四倍体の種は生息域を大きく広げています(図6)。この広い生息域には、アフリカツメガエルを含めた、幾つもの異質四倍体の種が生息していますが、それらの種はいずれも約1800万年前に一度だけ起きた異質四倍体化が基になっています。このように異質四倍体となった最初の種は、異なる2つのサブゲノムを獲得したことで、環境適応と生存競争に打ち勝つ進化の潜在能力が備わり、その結果、幾つもの種に進化しながら生息域を広げて行ったと考えられます。二倍体の祖先種が絶滅したのも、2つの種のそれぞれの優れた遺伝子をゲノムに合わせもった異質四倍体の子孫に凌駕されたため、と想像するに難くありません。
 |
|
図6. 異質倍数化により何がもたらされたか。アフリカにおける、ツメガエル属の二倍体種と異質四倍体種の生息域を示す(Evans et al, 2004改変)。二倍体種は赤道付近の熱帯地方に限られているが、四倍体種はチャドから南アフリカまで広く分布する。このことから、異質四倍体化により環境適応能力が増して広範囲に生活範囲を広げたと考えられる。なお、二倍体のネッタイツメガエルの適温は26度前後、異質四倍体のアフリカツメガエルは14~23度である。 |
全ゲノム重複は生物の進化の過程でしばしば起こる現象と考えられています。その例が、約5億年前の古生代カンブリア紀に脊椎動物が出現する過程で起きたとされる「2回の全ゲノム重複」です。これによって遺伝子数を格段に増やしたことが、脊椎動物の誕生とその後の多様化と繁栄をもたらした要因であったと考えられています。その後、脊椎動物の中には、さらに3回目、4回目の全ゲノム重複を起こしたものがいます。例えば、魚類の仲間の真骨魚類の系統では約3.2億年前に3回目の、さらにニジマスの系統では約1億年前に4回目の全ゲノム重複を起こしています(図1)。しかしいずれもゲノム重複後に1億年以上も経過しているため、サブゲノムを明らかにできていません。今回、約1800万年前という比較的最近に全ゲノム重複が起こったアフリカツメガエルのゲノムを解読することで、初めてサブゲノムを区別することができ、それを基に重複後のサブゲノムの変化を初めて明らかにすることができました。ツメガエル属のカエルの中でも、さらに4回目と5回目の全ゲノム重複が想定される種が見つかっています。今回のような解析をさらに進めることで、これまで謎であった約5億年前に起こったとされる脊椎動物の初期の進化での2回の全ゲノム重複や、約3.2億年前や約1億年前に起こったとされる魚類の系統での全ゲノム重複が、その後の進化にどのようなインパクトを与えたかを読み解く鍵、すなわちロゼッタストーンになるものと期待されます。このようにゲノムの中に痕跡として残されている脊椎動物の進化の道筋の謎を解き明かすことは、人類にとっての大きな知的財産となります。
全ゲノム情報の利用方法は多岐に渡ります。アフリカツメガエルはこれまでもモデル生物として、胚の発生や細胞の機能などにおける遺伝子の役割やその分子メカニズムの解析に使われてきましたが、今回の研究で得られた全ゲノム情報を用いることで、さらに多くの知見がもたらされると期待されます。例えば、遺伝子を改変する「ゲノム編集」という技術が近年注目されていますが、全ゲノム情報を基にこの技術を使えば、任意の遺伝子を改変してその遺伝子のもつ役割を解析することができます。アフリカツメガエルを用いたこれらの解析は、ヒトの遺伝的疾患の診断や治療などに役立つものであり、生命科学の発展に大きく貢献するものです。
◇本論文に関わった日本チームの機関と共著者一覧(18機関、23研究室)
東京大学
平良眞規(東京大学大学院理学系研究科生物科学専攻 准教授)
近藤真理子(東京大学大学院理学系研究科附属臨海実験所 准教授)
道上達男(東京大学 教授)
鈴木穣(東京大学大学院新領域創成科学研究科 教授)
国立遺伝学研究所
藤山秋佐夫(情報・システム研究機構国立遺伝学研究所先端ゲノミクス推進センター特任教授)
豊田敦(情報・システム研究機構国立遺伝学研究所生命情報研究センター 特任教授)
名古屋大学
松田洋一(名古屋大学大学院生命農学研究科応用分子生命科学専攻 教授)
宇野好宣(名古屋大学大学院生命農学研究科応用分子生命科学専攻 研究員)
広島大学
高橋秀治(広島大学大学院理学研究科両生類研究センター 特任准教授)
彦坂暁(広島大学大学院総合科学研究科 准教授)
鈴木厚(広島大学大学院理学研究科生物科学専攻・両生類研究センター 准教授)
基礎生物学研究所
上野直人(自然科学研究機構基礎生物学研究所形態形成研究部門 教授)
山本隆正(自然科学研究機構基礎生物学研究所形態形成研究部門 特任専門員)
高木知世(自然科学研究機構基礎生物学研究所形態形成研究部門 技術職員)
産業技術総合研究所
浅島誠(産業技術総合研究所創薬基盤研究部門 名誉フェロー)
原本悦和(産業技術総合研究所創薬基盤研究部門 主任研究員)
伊藤弓弦(産業技術総合研究所創薬基盤研究部門 研究グループ長)
北海道大学
福井彰雅(北海道大学大学院先端生命科学研究院 准教授)
長浜バイオ大学
荻野肇(長浜バイオ大学バイオサイエンス学部アニマルバイオサイエンス学科 教授)
山形大学
越智陽城(山形大学医学部メディカルサイエンス推進研究所 准教授)
国立成育医療研究センター
黒木陽子(国立成育医療研究センター ゲノム医療研究部 成育疾患ゲノム研究室 室長)
東京工業大学
田中利明(東京工業大学生命理工学院 助教)
徳島大学
渡部稔(徳島大学大学院総合科学研究部 教授)
立教大学
木下勉(立教大学理学部 教授)
メリーランド大学
太田裕子(メリーランド大学 助教)
北里大学
回渕修治(北里大学北里生命科学研究所 特任助教)
伊藤道彦(北里大学理学部 准教授)
バージニア大学
中山卓哉(バージニア大学 リサーチサイエンティスト)
新潟大学
井筒ゆみ(新潟大学 准教授)
沖縄科学技術大学院大学
安岡有理(沖縄科学技術大学院大学マリンゲノミックスユニット ポストドクトラルスカラー)
◇主な研究費
科研費新学術研究「ゲノム支援」(国立遺伝学研究所、東京大学、国立成育医療研究センター)、科研費・基盤(A、B、C)
◇その他の主な機関と主な共著者
カリフォルニア大学バークリー校(米国)(アダム・セッション、ダニエル・ロクサー、リチャード・ハーランド)
ウルサン国立科学技術研究所(韓国)(テジュン・クワン)
ラドバウンド分子生命科学研究所(オランダ)(サイモン・ファン・ヘーリンゲン、ガート・ヴィーンストラ)
ソーク研究所(米国)(イアン・キグレイ)
沖縄科学技術大学院大学(日本)(ダニエル・ロクサー、オレグ・シマコフ)